Research Article |
|
Corresponding author: Shu-Hong Li ( shuhongfungi@126.com ) Academic editor: Alfredo Vizzini
© 2023 Lin Li, Shan-Ping Wan, Yun Wang, Naritsada Thongklang, Song-Ming Tang, Zong-Long Luo, Shu-Hong Li.
This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Citation:
Li L, Wan S-P, Wang Y, Thongklang N, Tang S-M, Luo Z-L, Li S-H (2023) New species of Hydnotrya (Ascomycota, Pezizomycetes) from southwestern China with notes on morphological characteristics of 17 species of Hydnotrya. MycoKeys 100: 49-67. https://doi.org/10.3897/mycokeys.100.106709
|
Abstract
More specimens of Hydnotrya have been collected from southwestern China in recent years. Morphological and molecular analyses showed that they belonged to three species of Hydnotrya, of which two are new to science, H. oblongispora and H. zayuensis. The third one was H. laojunshanensis, previously reported in 2013. The new species are described, and their relationship to other species of Hydnotrya is discussed. H. laojunshanensis is re-described in more detail. The main morphological characters of 17 species of Hydnotrya are compared and a key to them is provided as well.
Key words
Discinaceae, hypogeous fungi, ITS, morphological diversity, taxonomy
Introduction
Hydnotrya Berk. & Broome is a genus of hypogeous fungi belonging to Pezizomycetes, Ascomycota. It was placed in the family Helvellaceae by
To date, nine Hydnotrya species have been reported in China: H. cerebriformis in Shanxi and Xinjiang, H. cubispora in Tibet, H. michaelis, H. tulasnei and H. brunneospora in Jilin (
Over the past two years, more Hydnotrya specimens have been collected in southwest China. Based on the morphological and molecular analyses, two new species were detected and described: H. oblongispora and H. zayuensis. Their relationships with other known Hydnotrya species are discussed and a more detailed supplementary description is given to another species H. laojunshanensis, previously found in Yunnan. Additionally, the main morphological characteristics of 15 species of Hydnotrya are listed and a key to the species of the genus is provided.
Materials and methods
The specimens were collected from Yunnan and Tibet, China. The type and other studied specimens were deposited at the Biological Science Museum of Dali University (BMDLU) and HKAS (Herbarium of Kunming Institute of Botany, Academy Sinica), China.
Descriptions of microscopic and macroscopic characters were based on specimens (BMDLU L20069, L20067, L21197, L21211, L21212, L21215, L21217, L22024, L22027, and HKAS95802) following the methods of
Total genomic DNA was extracted from the specimen using the OMEGA Plant Genomic DNA Kit. The internal transcribed spacer (ITS) rDNA region was amplified with PCR primers ITS1F and ITS4 (
ITS was used for the analysis of Hydnotrya species diversity in this study because ITS appears as a useful locus for the delimitation of Hydnotrya species. 46 ITS sequences from NCBI and this study representing 14 species of Hydnotrya (Table

Phylogeny derived from a maximum likelihood (ML) analysis of the nrDNA-ITS sequences from Hydnotrya species, using Gyromitra esculenta and G. infula as outgroup. Values next to nodes reflect, maximum likelihood bootstrap support values (BS), left, and Bayesian posterior probabilities (PP), right. Names of novel species and samples with newly generated sequences in bold. Symbols by taxon names indicate specific fruiting body types, the arrangement of the ascospores in the ascus and ascospore appearance.
Taxa information and GenBank accession numbers of the sequences used in this study. The newly generated sequences are in bold.
| Species name | Voucher | Origin | GenBank No. | Reference |
|---|---|---|---|---|
| Gyromitra esculenta | Gyr3 | France | AJ544208 |
|
| Gyromitra esculenta | m954 | UK | AJ544209 |
|
| Gyromitra infula | UBC F15196 | Canada | DQ384573 | GenBank |
| Gyromitra infula | Vellinga GLM | USA | AJ698480 |
|
| Hydnotrya badia | BJTC:FAN270 | China | NR_161070 | Yu et al. (2018) |
| Hydnotrya badia | BJTC:FAN270 | China | MH445399 | Yu et al. (2018) |
| Hydnotrya bailii | PRM 902032 | Czech | AM261522 | Stielow (2010) |
| Hydnotrya bailii | P.Reil_2 | Germany | GQ140239 | Stielow (2010) |
| Hydnotrya bailii | P.Reil | Germany | GQ140238 | Stielow (2010) |
| Hydnotrya bailii | 997 | Germany | GQ149465 | Stielow (2010) |
| Hydnotrya bailii | 979 | Germany | GQ149464 | Stielow (2010) |
| Hydnotrya brunneospora | HMAS 97138 | China | NR_161073 | Yu et al. (2018) |
| Hydnotrya brunneospora | HMAS 97138 | China | MH445404 | Yu et al. (2018) |
| Hydnotrya cerebriformis | 89_A12_Stielow | Germany | GQ140236 | Stielow (2010) |
| Hydnotrya cerebriformis | 87_G11_Stielow | Germany | GQ140235 | Stielow (2010) |
| Hydnotrya cerebriformis | BJTC:FAN647 | China | MH430537 | Yu et al. (2018) |
| Hydnotrya cerebriformis | GO-2010-097 | Mexico | KC152120 |
|
| Hydnotrya cerebriformis | GO-2009-455 | Mexico | KC152118 |
|
| Hydnotrya cerebriformis | GO-2009-242 | Mexico | KC152119 |
|
| Hydnotrya cubispora | SAT-13-273-01 | USA | MZ054357 | GenBank |
| Hydnotrya cubispora | K(M)104976 | UK | EU784273 |
|
| Hydnotrya laojunshanensis | YAAS L2425 | China | NR_132886 |
|
| Hydnotrya laojunshanensis | BMDLU L21211 | China | ON982580 | This study |
| Hydnotrya laojunshanensis | BMDLU L21212 | China | ON982593 | This study |
| Hydnotrya laojunshanensis | BMDLU L21215 | China | ON982594 | This study |
| Hydnotrya laojunshanensis | BMDLU L21197 | China | ON982592 | This study |
| Hydnotrya laojunshanensis | HKAS95802 | China | OP908303 | This study |
| Hydnotrya michaelis | K(M)61643 | UK | EU784275 |
|
| Hydnotrya michaelis | K(M)38647 | UK | EU784274 |
|
| Hydnotrya michaelis | 6463-307EMC | Germany | HM146816 |
|
| Hydnotrya nigricans | BJTC:FAN349 | China | NR_161071 | Yu et al. 2018 |
| Hydnotrya nigricans | BJTC:FAN349 | China | MH445400 | Yu et al. 2018 |
| Hydnotrya oblongispora | BMDLU L20067 | China | OM232075 | This study |
| Hydnotrya oblongispora | BMDLU L20069(Holotype) | China | OM232079 | This study |
| Hydnotrya oblongispora | BMDLU L21217 | China | OM232084 | This study |
| Hydnotrya puberula | BJTC:FAN721 | China | NR_161072 | Yu et al. 2018 |
| Hydnotrya puberula | BJTC:FAN721 | China | MH445401 | Yu et al. 2018 |
| Hydnotrya puberula | HMAS96758 | China | MH445402 | Yu et al. 2018 |
| Hydnotrya tulasnei | K(M)99871 | UK | EU784276 |
|
| Hydnotrya tulasnei | Berk. & Broome C34659 | Denmark | AJ969621 |
|
| Hydnotrya tulasnei | IT8 | Germany | GQ140240 | Stielow 2010 |
| Hydnotrya tulasnei | 605040 | Russia | KY401249 | GenBank |
| Hydnotrya variiformis | TK1615 | USA | AY558770 |
|
| Hydnotrya zayuensis | BMDLU L22024 | China | OP908304 | This study |
| Hydnotrya zayuensis | BMDLU L22027 (Holotype) | China | OP908305 | This study |
| Hydnotrya sp1. | SNF160 | USA | AY558768 |
|
| Hydnotrya sp2. | SNF82 | USA | AY558769 |
|
| Hydnotrya sp3. | JT19176 | USA | MN653030 | GenBank |
| Hydnotrya sp4. | JT19085 | USA | MN653044 | GenBank |
| Hydnotrya sp5. | JLF2015 | USA | MH220061 | GenBank |
Results
Phylogenetic analysis
The ML and Bayesian analyses of the 50 ITS sequences, are shown in Fig.
In the phylogenetic tree, the 46 ITS sequences from Hydnotrya ascomata revealed the phylogenetic relationship of 14 species: Clade 1 includes 5 sequences of H. bailii from Europe. Clade 2 includes 2 sequences of H. brunneospora from China. Clade 3 includes 4 sequences of H. tulasnei from Europe. Clade 4 includes 3 sequences of H. puberula from China. Clade 5 includes 2 sequences of H. badia from China. Clade 6 includes 2 sequences of H. nigricans from China. Clade 7 includes 6 sequences of H. cerebriformis from Germany, China, and Mexico; two other distinct clades were revealed, one comprising Eurasian specimens, and the other comprising specimens from Mexico, which is probably because these specimens, respectively, are from Holarctic and Neotropical regions. Clade 8 includes 3 sequences of H. variiformis from the USA. Clade 9 includes 2 sequences of H. cubispora from the UK and USA. Clade 10 includes 3 sequences of H. michaelis from Europe. Clade 11 includes 3 sequences of new species, H. oblongispora from China. Clade 12 includes 3 sequences of Hydnotrya sp. from the USA. They may be new species from North America that have not yet been reported. Clade 13 includes 6 sequences of H. laojunshanensis from China. When the latter was reported, only one specimen was found, and many more were collected over the past few years, so new DNA sequences of H. laojunshanensis were added. Clade 14 includes 2 sequences of a new species, H. zayuensis from China. The phylogenetic analysis shows that the new species are distinct from other Hydnotrya species. In addition to the ITS sequences used in this phylogenetic analysis, the LSU sequences were amplified from the newly supplemented specimens in this study and uploaded to NCBI for future study.
Based on the ITS locus, two major monophyletic lineages are presented, showing a strong sister relationship (BS=100%; PP = 1.0). They are Clade A (including Clade 1–9) and Clade B (include Clade 10–14) respectively. The species included in these two phylogenetic morphologically share commonalities and uniqueness.
Taxonomy
Hydnotrya oblongispora
Diagnosis
Differs from other species in the genus Hydnotrya by its nearly single-chambered ascomata and long ellipsoidal ascospores.
Etymology
oblongispora, refers to the long ellipsoidal ascospores.
Holotype
China, Yunnan, Lijiang (26°37.00'N, 99°42.00'E), alt. 3737 m, in the forest of Abies forrestii Coltm.-Rog, 12 August 2020, Lin Li, BMDLU L20069.
Description
Ascomata irregularly globose, 1.0–2.5 cm in diameter when fresh, smooth, sometimes gently folded inward, surface light khaki (4C5) to reddish brown (8D8); nearly single-chambered with a primary apical opening up to 0.2–0.8 cm in diameter, sometimes the opening is just an almost closed seam, white fluffy inside cavity. Elastic and crisp. No special smell was noticed.
Peridium two-layered, 280–340 µm thick, outer layer 80–100 µm thick, composed of light brown (6D8) ellipsoidal or irregular cells, with a red brown (6E8) pigment deposited on the outermost cells; inner layer, 200–240 µm thick, consists of hyaline interwoven hyphae. Gleba chamber hollow, lined with a milky white (4B2) hymenium, hymenial surface fluffy. Asci cylindrical, 102.5–138.5 × 13.0–25.5 µm, 8-spored, thin-walled, narrowed into a long stalk (20–35 μm) at the base, without croziers, arranged in a palisade. Ascospore strictly uniseriate, long ellipsoidal, (20.0–) 26.5–39.0 × (9.5–) 11.0–21.5 μm, Q = 2.0±0.03, hyaline when immature, golden yellow (5B7) when mature, with a thickened exosporium, surface pitted. Paraphyses hyaline, straight stick shape, 2.5–5 µm in diam, septate, exceeding the asci by 60–70 µm.
Ecology and distribution
Hypogeous, solitary, or in groups in soil, under A. forrestii mixed with shrubs of Rhododendron spp., fruiting from late summer to early autumn. Known only from Yunnan Province, China.
Additional specimens examined
Notes
H. oblongispora is characterized by its mostly simple-chambered ascomata and golden yellow long-ellipsoid ascospores, especially with pitted surfaces, which differ from all other species of Hydnotrya. Molecular analysis also shows that H. oblongispora is distinct from other Hydnotrya species, although it is closely related to H. michaelis. However, H. michaelis has convoluted, lobed ascomata and broadly ellipsoid spores with warty ascospores, which differ from this new species.

Hydnotrya oblongispora A young sarcomata B mature ascomata with different openings C a piece of section of the ascomata in lactophenol cotton blue D a peridium section in lactophenol cotton blue E a section of paraphyses in 5% KOH F a base of asci in lactophenol cotton blue G ascospores released from the ascus H asci in lactophenol cotton blue I an ascus with 8 ascospores J–L ascospores under SEM. Scale bars: 1 cm (A, B); 100 μm (C); 50 μm (D); 10 μm (E–I); 5 μm (J, L); 2 μm (K).
Hydnotrya zayuensis
Diagnosis
Differs from all other species in Hydnotrya by its almost single-chambered ascomata, light golden yellow ellipsoidal ascospores.
Etomology
zayuensis from Latin, referring to the type locality.
Holotype
China, Tibet, Zayu (28°35.00'N, 98°06.00'E), alt. 3770 m, in a forest of Abies sp., 11 August 2022, Lin Li BMDLU L22027.
Description
Ascomata irregularly globose, 1.5–2.5cm in diameter when fresh, smooth, convoluted, almost single-chambered with a primary apical opening, sometimes the opening nearly closed like a seam, white fluffy inside, surface cinnamon (5E8); shrunken, becoming fuzzy when dried, although there are no protruding hyphae cells from the outermost layer of the peridium. Elastic to crisp. No special smell was noticed.
Peridium two-layered, 180–250 µm thick, outer layer 40–80 µm thick, composed of ellipsoid or irregular cells, which grow larger toward the surface, with a yellow brown (4C5) pigment deposited on the outermost cells; inner layer, 110–160 µm thick, consisting of hyaline parallel interwoven hyphae. Gleba chamber hollow, lined with off-white (1A2) hymenium when immature; two-layered when mature, the outer layer golden brown (5C7), the inner layer yellowish to whitish (4A2), hymenial surface fluffy. Asci cylindrical, 118.5–130.5 × 15.0–22.5 µm, 8-spored, thin-walled, narrowed into a long stalk (20–40 μm) at the base, without croziers, arranged in a palisade. Ascospore strictly uniseriate, ellipsoid (shape including the thickened exosporium), (17–)20–30.5 × 15.5–18.0 μm, Q = 1.5 ± 0.16, hyaline, exosporium thin when immature, surface roughness, and looks crumbly, golden yellow (4B8) when mature. Paraphyses hyaline, straight stick shape, 1.5–2.5 µm in diam, septate, apical slightly inflated, exceeding the asci by 120–160 µm.

Hydnotrya zayuensis A ascomata B section of ascomata, with hymenium-lined chambers C habitat D inner surface of ascomata E peridium in 5% KOH F hymenium G asci in 5% KOH H paraphyses I ascospores in 5% KOH J–L ascospores under SEM (L. SEM of a single ascospore cut in half). Scale bars: 1cm (A); 1 mm (B); 0.5cm (D); 100 μm (E); 50 μm (F); 20 μm (G); 10 μm (H); 10 μm (I); 5 μm (J–L).
Ecology and distribution
Hypogeous, solitary in the humus under Abies sp. mixed with shrubs of Rhododendron spp. Fruiting in summer, from July to September. Known only from Zayu, Tibet, China.
Additional specimen examined
Notes
Morphologically, H. zayuensis is similar to H. laojunshanensis. However, H. zayuensis has much smaller ascospores, and a thinner peridium, as well as lighter colored ascomata. Molecular analysis showed that H. zayuensis is distinct from H. laojunshanensis and other species of Hydnotrya.
Hydnotrya laojunshanensis
Description
Ascomata irregularly globose, 1.0–3.0 cm in diameter when fresh, brownish orange (6C8), smooth, mostly single-chambered with a primary apical opening to 0.1–0.5 cm in diameter, the opening rarely narrowing into a slit, sometimes folded forming few channels, lined with white fluffy hymenium. Elastic to crisp. No special smell was noticed.
Peridium two-layered, 350–570 µm thick, outer layer 160–200 µm thick, composed of light brown (6E8) angular or irregular cells, inner layer, 220–350 µm thick, consisting of hyaline interwoven hyphae. Gleba chamber hollow, lined with off-white (1A2) hymenium when immature; two-layered when mature, the outer layer orange (6B8), the inner layer yellowish to whitish (4A2), hymenial surface fluffy. Asci cylindrical, 331.5–390.5 × 25.5–35.5 µm, 8-spored, thin-walled, narrowed at the base into a long stalk (30–50 μm), without croziers, arranged in a palisade. Ascospore strictly uniseriate, ellipsoid (excluding the thickened exosporium), rectangular (with the exosporium), (26.5–)33.0–50.5 × (15.5–)20.5–35.5(–38.0) µm Q = 1.35±0.02, surface rough, reddish orange to golden (6B8) when mature. Paraphyses hyaline, straight stick shape, 2.0–6 µm in diam, apical slightly inflated, septate, exceeding the asci by 180–300 µm.

Hydnotrya laojunshanensis A young sarcomata cut in half B mature ascomata with one cut in half C infolded and chambered ascoma D section of hymenium in 5% KOH E a peridium section in 5% KOH F ascospores released from asci in 5% KOH G–I ascospores under SEM (I. SEM of a single ascospore cut in half). Scale bars: 1 cm (A, B); 50 μm (D, E); 20 μm (F); 5 μm (G–I).
Ecology and distribution
Hypogeous, solitary, or in groups in soil, under Abies spp., fruiting from late summer to early autumn. Known only from Yunnan Province, China.
Additional specimens examined
China, Yunnan Province, Laojun mountains, 26°42.00'N, 99°42.00'E, alt. 3786 m, in a forest of A. forrestii var. smithii, 30.Aug.2012, Lin Li (Holotype, YAAS L2425; GenBank KC878618); Shangri-La, 28°16.00'N, 99°11.00'E, alt. 3978 m, in a forest of Abies sp., 19 Aug. 2014, Shanping Wan (HKAS95802 GenBank: ITS = OP908303), Lijiang, 26°42.00'N, 99°58.00'E, alt. 3540 m, in a forest of A. forrestii, 12 Sept. 2019, Lin Li (BMDLU L21197 GenBank: ITS = ON982592, LSU = ON982620); Lijiang, 26°56.00'N, 99°32.00'E, alt. 3805 m, in a forest of A. forrestii, 21 Sept. 2021, Lin Li (BMDLU L21211 GenBank: ITS = ON982580, LSU = ON982621, BMDLU L21212 GenBank: ITS = ON982593, LSU = ON982622, BMDLU L21215 GenBank: ITS = ON982594, LSU = ON982623).
Notes
When the species was described in 2013 by Li et al., only one collection from Mt. Laojun in Yunnan Province, China, was reported. More specimens of H. laojunshanensis have been found at other places in Yunnan since then. We discovered that this species had not only simple chambered ascomata but also folded, chambered ascomata. This species has large, rectangular ascospores (including thickened exsporium) with a rough surface differentiating from other species in Hydnotrya.
Discussion
To date, 17 species of Hydnotrya (including these two new species) are accepted worldwide (
| Species | Ascomata | Gleba | Ascospore | Asci | Host Plants | Distribution | References | |
|---|---|---|---|---|---|---|---|---|
| Hydnotrya badia L. Fan, Y.W. Wang & Y.Y. Xu 2018 | Irregularly subglobose, 7–15 × 14–19 mm diam., surface even, brown to earth brown. | Gleba solid, with numerous variably compacted canals and chambers (usually without empty space). |
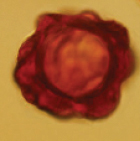
|
Roughly globose, 25–40 μm in diam. including ornamentation, 17.5–27.5 μm in diam. excluding ornamentation, red-brown to reddish, thickened exisporium with regular large protuberances | Asci broadly clavate to somewhat saccate, sessile, or narrowed at the base into a short stalk, 125–172.5 × 65–75 μm, randomly immersed in paraphyses, 8-spored, spores mostly biseriate. | Pinus sp. | Huize, China Asia 2000–2900m | Yu et al. 2018 |
| Hydnotrya bailii Soehner 1959 | Irregularly subglobose, 10–20(–25) mm diam., dark brown, with deep furrows often with multiple lobes, with one or many openings at the apex, with pleasant aromatic smell. | Gleba solid, dark brown, strongly convoluted cavities. |
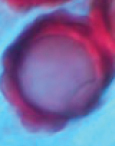
|
Globose, (27.5-) 30–34 (–37.5) μm in diam, brown reddish, exosporium thickening with blistered warts | Asci cylindrical, 250–300×30–40 μm, 8-spored, spores mostly uniseriate. | Picea abies | Europe |
|
| Hydnotrya brunneospora L. Fan, Y.W. Wang & Y.Y. Xu 2018 | Irregularly globose, 20–23 mm diam., dark brown when dry, surface smooth. | Gleba solid, scattered with some small, isolated, and irregularly shaped chambers. |
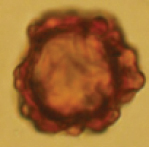
|
Roughly globose, 26.25–46.25 μm diam., brown to golden brown at maturity (never reddish), exosporium thickening with small protuberances. | Asci cylindrical to clavate, narrowed at the base into a short stalk, 162.5–237.5 × 30–47.5 μm, randomly immersed in paraphyses, 8-spored, spores mostly uniseriate. | Betula platyphylla | Jilin, China Asia | Yu et al. 2018 |
| Hydnotrya cerebriformis Harkn. 1899 | Irregular spherical, lobulated, 10–35 × 10–20 mm diam., reddish-brown, cerebriform, with cavities that communicate with the gleba. | Gleba with labyrinthine chambers composed of invagination and fusion from the walls of the ascoma. |
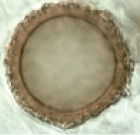
|
Globose ascospores 20–25μm diam. (x = 22.85 μm), excluding ornamentation, amber-brown, walls 1μm wide. Finely warty ornamentation, warts up to 4μm long. | Asci cylindrical, 175–200 × 25–35 μm, 8-spored, spores mostly uniseriate. | Pinus sp. Abies sp. | Europe North America 3100–4000m |
|
|
Hydnotrya confusa |
Ovoid or irregular, size from ca. 20 × 20 × 15 cm up to 40 × 40 × 20 mm, greyish-brown or red-brown, with a primary apical opening and sometimes some smaller secondary openings. | Gleba hollow, with single chambered but mostly cerebriform folded. |
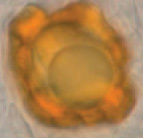
|
Ovoid or very broad ellipsoid, 38–50 × 28–32 µm, golden brown, exosporium much thickened, vertically grooved, forming irregular warts. | Asci cylindrical, 290–320 × 38–43 μm, 8-spored, spores mostly uniseriate. clavate at immature, with irregular or biseriate, cylindric at maturity, strictly uniseriate. | Picea sp. | Europe 361m |
|
|
Hydnotrya cubispora (E.A. Bessey & B.E. Thomps.) |
Irregularly globose, 5–10 mm diam., Isabella color, with somewhat cerebriform folds radiating distinctly from central opening | Gleba with cavity simple, but somewhat irregular due to surface lobing. |
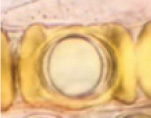
|
Cubical, 47–50 × 23–32µm, including thickening exosporium, brownish, with maturity. | Asci cylindrical, 100–120 μm long, 8-spored, spores mostly uniseriate. | Coniferous forest | Europe North America |
|
| Hydnotrya inordinata Trappe & Castellano, 2000 | Irregular globose, 8–30mm diam., dark red-brown, convolute and infolded ptychothecia with one or a few openings from the interior | Gleba complex, of infolded tramal plates forming canals and chambers 0.5–3mm broad. |

|
Globose to ellipsoid, 20–30×20–28μm excluding ornamentation, brown-yellow, with aggregated , irregular flexuous spines | Asci cylindrical, ±300 × 25–33 μm, (6–) 8-spored, spores mostly uniseriate. | Abies amabilis Tsuga mertensiana | North America 1800m |
|
| Hydnotrya laojunshanensis Lin Li, D.Q. Zhou & Y.C. Zhao 2013 | Irregularly globose, 10–30 mm diam., brownish orange, smooth, mostly single-chambered with a primary apical opening, rare the opening narrowing into a slit, sometimes folded forming a few channels, lined with white fluffy hymenium. No special smell. | Gleba hollow, single-chambered, sometimes infolded and chambered, lined with hymenium with orange asci and whitish to yellowish paraphyses. |
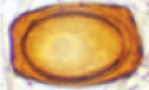
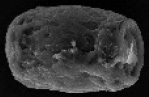
|
Ellipsoid without thickened exosporium, rectangle (including exosporium), (42.5–) 50.0–57.2(–60.3) × (27.5–)30.4–36.9(–38.2) µm, reddish orange, thickening exosporium with rough surface. | Asci cylindrical, 331.5–390.5 × 25.5–35.5 µm, 8-spored, spores strictly uniseriate | Abies spp. | Yunnan, China Asia 3500–3800m |
|
|
Hydnotrya michaelis (E. Fisch.) |
Irregular or subspherical, up to 60 mm across, with rounded opening, wrinkled, lobulate, with numerous invaginations, odor very strong, somewhat pungent, rather persistent. | Gleba labyrinthoid, with large, sinuous cavities, separated by folded inwards portions of ascoma wall. |
|
Broadly ellipsoid, (21.2–)24.9–29.6(–32.2) ×(18.8–)19.8–22.4 (–24.9) µm; ornamentation excluded), honey-yellow, exosporium thickened, with conspicuous, irregular, often interconnected warts | Asci cylindrical, 200–220 × 30–35 µm, 8-spored, spores strictly uniseriate | Pinaceae | Europe North America |
|
| Hydnotrya nigricans L. Fan, Y.W. Wang & Y.Y. Xu 2018 | Irregular globose, 13 × 9mm, black brown to blackish | Gleba solid, brown, red to dark reddish, with some irregularly shaped and isolated small chambers lined with pale whitish hymenium. |
|
Irregularly globose, 25.0–37.5 μm in diam., red brown, exosporium unevenly thickened, and usually of trigonal outline in cross section | Asci broadly clavate to saccate, sessile or narrowed at the base into a short stalk, 87.5–190 × 25–62.5 μm, scattered between paraphyses in a hymenium,8-spored,with spores mostly biseriate. | Pinus sp. | Sichuan, China Asia | Yu et al. 2018 |
| Hydnotrya oblongispora sp. nov. | Irregularly globose, 10–25mm in diam. when fresh, light khaki to reddish brown, smooth, mostly single-chambered with a primary apical opening up to 02–08 mm in diam., sometimes infolded. | Gleba hollow, single-chambered lined with milky white hymenium, hymenium surface fluffy. |


|
Long-ellipsoid (20.0–) 26.5–39.0 × (9.5–) 11.0–21.5 μm, golden brown, thickened exporium with pitted surface. | Asci cylindrical, 102.5–138.5 × 13.0–25.5 µm, narrowed at the base into a long stalk (20–30 μm), 8-spored, spores strictly uniseriate | Abies forrestii | Yunnan, China Asia 3500–4000m | This study |
| Hydnotrya puberula L. Fan, Y.W. Wang & Y.Y. Xu 2018 | Irregularly subglobose, 11–20 × 8–19 mm, brown to dark brown, sometimes with purple tints when fresh, much convoluted with deep furrows, ascoma surface tomentulose | Gleba solid, compact, dark brown to purple reddish at maturity, with numerous small chambers. |
|
Roughly globose, 22.5–42.5 μm in diam., red brown to reddish, exosporium unevenly thickened by irregularly large protuberances. | Asci clavate to saccate, 125–190 × 55–80 μm, sessile or with a short stalk, borne among palisade-like paraphyses in the hymenium, 8-spored, with spores mostly biseriate. | Pinus sp. | Yunnan, China Asia | Yu et al. 2018 |
| Hydnotrya soehneri Svrček, 1955 | Irregularly subglobose, tuberous, 10–40 mm wide, reddish and reddish-gray to reddish brown, odor light fragrance | Gleba solid, whitish to yellowish gray, at maturity is colored reddish-brown corridors (from mature spores). | Spherical, 25 – 36 (– 42), red brown, exosporium thickened, coarsely warty. | Asci mostly cylindrical to saccate, 150–300 × 35–70 µm, 8-spored, mostly incompletely arranged biseriate. | Mixed woods | Europe |
|
|
| Hydnotrya subnix Trappe & Castellano, 2000 | Irregular subglobose, 50–65mm in diam, dark red-brown, glabrous to minutely roughened. Odor and taste strongly of spicy garlic. | Gleba variable, deeply convoluted and infolded lacking openings from the interior, forming canals and locules 1–10mm broad. |
|
Globose to rarely ellipsoid, 23–30μm in diam. excluding ornamentation, brown, coarsely warty. | Asci mostly cylindrical, 300–340 × 25–40 µm, 8-spored, mostly incompletely arranged uniseriate | Abies amabilis | North America 950m |
|
| Hydnotrya tulasnei (Berk.) Berk. & Broome, 1846 | Irregularly spherical or lobed, sometimes with inward folds, 20–70 mm diam., ochre-reddish to brick red | Gleba solid, later yellow brown, with labyrinthic chambers. |
|
Globose, 20–30 (–33) μm diam. (including ornamentation), ochre-reddish, with conspicuous, irregular warts. | Asci broadly clavate or cylindrical, 175–210 × 30–62.5 µm, (4–) 8-spored, spores biseriate. | Coniferous forest | Europe North America 1600m |
|
| Hydnotrya variiformis Gilkey, 1947 | Globose to subglobose to flattened, somewhat depressed, 7–40 mm broad, cinnamon-buff to cream-buff | Gleba variable, from a simple cavity to extremely lobed with numerous small chambers the interior, usually opening to the exterior at one or more points. |
|
Ellipsoid, 24–28×36–36 μm, yellow-brown, thickened exosporium wall, surface appearing punctate and with small irregular nodules | Asci 240–280 × 24 µm, 8-spored, clavate at immaturity, spores incompletely biseriate; cylindrical at maturity, spores strictly uniseriate. | Coniferous forest | North America 1200–2400m |
|
| Hydnotrya zayuensis sp. nov. | Irregularly globose, 15–20 mm in diameter when fresh, smooth, gentle inward folds, surface cinnamon. Mostly single-chambered with a primary apical opening, the opening is just an almost closed seam, white fluffy inside cavity. Elastic and crisp. No special smell. | Gleba hollow, single-chambered with a primary apical opening, sometimes the opening is just an almost closed seam. |

|
Ellipsoid, (17–)20–30.5 × 15.5–18.0 μm, (including thickened exosporium), golden yellow, surface rough, looking like crumbly. | Asci cylindrical, 118.5–130.5×15.0–22.5 µm, 8-spored, spores strictly uniseriate | Abies sp. | Zayu, China Asia 3770m | This study |
The ascospore morphology is highly variable among different species in Hydnotrya, which is useful for distinguishing species.
Based on ITS analyses, 14 species of Hydnotrya are divided into two lineages, A and B. The species in the clade A mostly have nearly solid gleba (6 out of 9) and globose, warty ascospores, either uniseriately or biseriately arranged in asci. The clade A is divided into two subclades: the subclade Aa (clade 1–6) and Ab(clade 7–9). The species in the subclade Aa have solid ascomata. Two groups can be distinguished: the group 1 (clade 1 and 2) and group 2 (clade 3–6), both found in China and Europe. The group 1 contains two species with ascospores uniseriately arranged in asci; the group 2 contains four species with ascospores biseriately arranged in asci. Species in the subclade Ab are distributed in China, Europe, and America, and have hollow ascomata and ascospores uniseriately arranged in asci. The species in the clade B has hollow to chambered gleba and ellipsoidal ascospores (without thickened exosporium), biseriately arranged in asci. The clade B is divided into two groups: Ba and Bb. The group Ba (clade 10 and 11) contains 2 species distributed in China and Europe, with ellipsoidal ascospores, with a pitted surface. The group Bb (clade 13 and 14) contains two species, only found in China, with rectangular and ellipsoidal ascospores (with thickened exosporium), with a rough surface. (Fig.
Based on the morphological and molecular phylogenetic analyses there seems to be a trend in morphological traits among the species within the genus Hydnotrya, that is, the gleba evolved from being hollow or chambered to nearly solid; the ascus becoming shorter and wider, with ascospores arranged from uniseriate to biseriate; ascospores from ellipsoidal to globose, with an ornamentation from smooth to rough as well. This evolutionary trend in the genus Hydnotrya is probably related to their hypogeous habits, that is, if the gleba has more chambers, the ascoma will hold more ascospores, and so there are more chances of ascospores to be dispersed by animals that eat them (
In China, 9 species were recorded before this study (
Key to species of Hydnotrya
| 1 | Ascomata hollow, gleba chamber simple or infolded | 2 |
| – | Ascomata solid, gleba labyrinthine chambered | 11 |
| 2 | Ascospores rectangular or cubical | 3 |
| – | Ascospores ellipsoidal or globose | 4 |
| 3 | Ascospores cubical | H. cubispora |
| – | Ascospores rectangular | H. laojunshanensis |
| 4 | Odor distinct, with a special smell | 5 |
| – | Odor not distinct | 6 |
| 5 | Odor and taste strongly garlic | H. subnix |
| – | Odor strong pungent and persistent | H. michaelis |
| 6 | Ascospores mostly globose | 7 |
| – | Ascospores ellipsoidal or long ellipsoidal | 8 |
| 7 | Ascospores globose, with prominent echinate ornamentation | H. cerebriformis |
| – | Ascospore mostly globose, with aggregated, irregular flexuous spines | H. inordinata |
| 8 | Ascospores long ellipsoidal, surface pitted, ascomata mostly single chambered | H. oblongispora |
| – | Ascospores ellipsoidal, Q ratio less than 2 | 9 |
| 9 | Ascospores incompletely biseriate at immaturity, strictly uniseriate at maturity in asci | 10 |
| – | Ascospores strictly uniseriate from immature to mature asci | H. zayuensis |
| 10 | Ascospores broadly ellipsoidal, vertically grooved, forming irregular warts | H. confusa |
| – | Ascospores ellipsoidal, surface appearing punctate and with small irregular nodules | H. variiformis |
| 11 | Ascospores mostly uniseriate | 12 |
| – | Ascospores mostly biseriate | 13 |
| 12 | Ascospores less than 35 μm |
H. bailii |
| – | Ascospores up to 46 μm* in length, brown to golden brown | H. brunneospora |
| 13 | Odor with a light fragrance | H. soehneri |
| – | Odor not distinct | 14 |
| 14 | Ascoma surface tomentose, withpurple tints when fresh | H. puberula |
| – | Ascoma not tomentose | 15 |
| 15 | Ascospores without prominent protuberances, trigonal outline in cross section, ascomata blackish | H. nigricans |
| – | Ascospores with recognizable protuberances | 16 |
| 16 | Ascospores, 20–30 μm diam.*, ochre-reddish, with conspicuous, irregular warts | H. tulasnei |
| – | Ascospores, 25–40 μm in diam.*, red brown to reddish, with regular large protuberances | H. badia |
Acknowledgments
Thanks to Mr. Shucheng He for his help in specimen collection. Thanks to the Fungal Diversity Conservation and Utilization Team in Northwest Yunnan for providing the research platform and team members for their help.
Additional information
Conflict of interest
The authors have declared that no competing interests exist.
Ethical statement
No ethical statement was reported.
Funding
This study was financially supported by the National Natural Science Foundation of China (No. 31800009, 32060008) and the Yunnan Fundamental Research Project (2017FD135).
Author contributions
Data curation: SPW. Methodology: SMT. Writing - original draft: LL. Writing - review and editing: YW, NT, SHL, ZLL.
Author ORCIDs
Lin Li https://orcid.org/0009-0000-8167-2965
Shan-Ping Wan https://orcid.org/0000-0002-0794-3701
Naritsada Thongklang https://orcid.org/0000-0001-9337-5001
Song-Ming Tang https://orcid.org/0000-0002-6174-7314
Zong-Long Luo https://orcid.org/0000-0001-7307-4885
Shu-Hong Li https://orcid.org/0000-0001-5806-9148
Data availability
All of the data that support the findings of this study are available in the main text.
References
- Abbott SP, Currah RS (1997) The Helvellaceae systematic revision and occurrence in northern and northwestern North America. Mycotaxon 62: 1–125.
- Bemmann M, Bandini D (2011) A collection of Hydnotrya confusa Spooner 1992 from Southwest Germany. Ascomycete. Org 3(3): 55–60.
- Beug M, Bessette AE, Bessette AR (2014) Ascomycete fungi of North America: a mushroom reference guide. University of Texas Press. New York, USA, 16–18. https://doi.org/10.7560/754522
- Bonito G, Smith ME, Nowak M, Healy RA, Guevara G, Cázares E, Kinoshita A, Nouhra ER, Domínguez LS, Tedersoo L, Murat C, Wang Y, Moreno BA, Pfister DH, Nara K, Zambonelli A, Trappe JM, Vilgalys R (2013) Historical biogeography and diversification of truffles in the Tuberaceae and their newly identified southern hemisphere sister lineage. PLoS ONE 8(1): e52765. https://doi.org/10.1371/journal.pone.0052765
- Brock PM, Döring H, Bidartondo MI (2009) How to know unknown fungi: The role of a herbarium. The New Phytologist 181(3): 719–724. https://doi.org/10.1111/j.1469-8137.2008.02703.x
- Bryan L (2003) Hydnotrya cubispora in Wales. Field Mycology 4(4): 122. https://doi.org/10.1016/S1468-1641(10)60218-6
- Burdsall-Jr HH (1968) A revision of the genus Hydnocystis (Tuberales) and of the hypogeous species of Geopora (Pezizales). Mycologia 60(3): 496–525. https://doi.org/10.1080/00275514.1968.12018600
- Cox F, Barsoum N, Lilleskov EA, Bidartondo MI (2010) Nitrogen availability is a primary determinant of conifer mycorrhizas across complex environmental gradients. Ecology Letters 13(9): 1103–1113. https://doi.org/10.1111/j.1461-0248.2010.01494.x
- Dimitrova E, Gyosheva M (2008) Hypogeous ascomycetes in Bulgaria. Phytologia Balcanica 14(3): 309–314.
- Gardes M, Bruns TD (1993) ITS primers with enhanced specificity for basidiomycetes application to the identification of mycorrhizae and rusts. Molecular Ecology 2(2): 113–118. https://doi.org/10.1111/j.1365-294X.1993.tb00005.x
- Gilkey HM (1939) Tuberales of North America. Oregon State Monographs. Studies in Botany 1: 1–63.
- Gilkey HM (1947) New or otherwise noteworthy species of Tuberales. Mycologia 39(4): 441–452. https://doi.org/10.1080/00275514.1947.12017626
- Gilkey HM (1954) Taxonomic notes on Tuberales. Mycologia 46(6): 783–793. https://doi.org/10.1080/00275514.1954.12024414
- Hall T (2007) BioEdit 7.0. 5.3 Department of Microbiology, North Carolina State University. http://www.mbio.ncsu.edu/BioEdit/Bioedit.html
- Hansen K, Pfister DH (2006) Systematics of the Pezizomycetes–The operculate discomycetes. Mycologia 98(6): 1029–1040. https://doi.org/10.3852/mycologia.98.6.1029
- Harkness HW (1899) Californian hypogeous fungi. Proceedings of the California Academy of Sciences 3: 266.
- Hawker LE (1955) Hypogeous fungi. Biological Reviews of the Cambridge Philosophical Society 30(2): 127–158. https://doi.org/10.1111/j.1469-185X.1955.tb01578.x
- Izzo AD, Meyer M, Trappe JM, North M, Bruns TD (2005) Hypogeous ectomycorrhizal fungal species on roots and in small mammal diet in a mixed-conifer forest. Forest Science 51(3): 243–254.
- Katoh K, Standley DM (2013) MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Molecular Biology and Evolution 30(4): 772–780. https://doi.org/10.1093/molbev/mst010
- Kellner H, Luis P, Buscot F (2007) Diversity of laccase-like multicopper oxidase genes in Morchellaceae: Identification of genes potentially involved in extracellular activities related to plant litter decay. FEMS Microbiology Ecology 61(1): 153–163. https://doi.org/10.1111/j.1574-6941.2007.00322.x
- Kirk PM, Cannon PF, Minter DW, Stalpers JA (2008) Ainsworth & Bisby’s dictionary of the fungi, 10th edn. CAB International, Wallingford, 325 pp. https://doi.org/10.1079/9780851998268.0000
- Kornerup A, Wanscher JH (1978) Methuen Handbook of Colour (3rd edn.). Methuen, London, 252 pp.
- Kumar LM, Smith ME, Nouhra ER, Orihara T, Leiva PS, Pfister DH, McLaughlin DJ, Trappe JM, Healy RA (2017) A molecular and morphological re-examination of the generic limits of truffles in the Tarzetta-Geopyxis lineage–Densocarpa, Hydnocystis, and Paurocotylis. Fungal Biology 121(3): 264–284. https://doi.org/10.1016/j.funbio.2016.12.004
- Læssøe T, Hansen K (2007) Truffle trouble: What happened to the Tuberales? Mycological Research 111(9): 1075–1099. https://doi.org/10.1016/j.mycres.2007.08.004
- Ławrynowicz M (1990) Chorology of the European hypogeous Ascomycetes. II. Acta Mycologica 26(1): 7–75. https://doi.org/10.5586/am.1990.001
- Li L, Zhao YC, Zhang XL, Su HY, Li SH, Zhou DQ (2013) Hydnotrya laojunshanensis sp. nov. from China. Mycotaxon 125(1): 277–282. https://doi.org/10.5248/125.277
- Miller MA, Pfeiffer W, Schwartz T (2011) The CIPRES science gateway: a community resource for phylogenetic analyses. Proceedings of the 2011 TeraGrid Conference: extreme digital discovery. New York, NY, United States, 18 July 2011. Association for Computing Machinery, 1–8. https://doi.org/10.1145/2016741.2016785
- Nylander JA, Ronquist F, Huelsenbeck JP, Nieves-Aldrey J (2004) Bayesian phylogenetic analysis of combined data. Systematic Biology 53(1): 47–67. https://doi.org/10.1080/10635150490264699
- O’Donnell K, Cigelnik E, Weber NS, Trappe JM (1997) Phylogenetic relationships among ascomycetous truffles and the true and false morels inferred from 18S and 28S ribosomal DNA sequence analysis. Mycologia 89(1): 48–65. https://doi.org/10.1080/00275514.1997.12026754
- Piña-Páez C, Garibay-Orijel R, Guevara-Guerrero G, Castellano MA (2017) Descripción y distribución de Hydnotrya cerebriformis (Discinaceae: Pezizales) en México. Revista Mexicana de Biodiversidad 88(2): 269–274. https://doi.org/10.1016/j.rmb.2017.03.017
- Rambaut A, Drummond AJ, Xie D, Baele G, Suchard MA (2018) Posterior summarization in Bayesian phylogenetics using Tracer 1.7. Systematic Biology 67(5): 901–904. https://doi.org/10.1093/sysbio/syy032
- Ronquist F, Huelsenbeck J, Teslenko M (2011) Draft MrBayes version 3.2 manual: tutorials and model summaries. [Distributed with the software] http://brahms.biology.rochester.edu/software.html
- Slavova M, Assyov B, Denchev TT, Denchev CM (2021) Hydnotrya michaelis–an Uncommon Fungus from Unexpected Habitat. Ecologia Balkanica 13(1): 167–171.
- Soehner E (1942) Tuberaceen-Studien. Notizblatt des Botanischen Gartens und Museums zu Berlin-Dahlem 15(5): 762–782. https://doi.org/10.2307/3995172
- Spooner BM (1992) A new species of Hydnotrya (Helvellaceae) from the British Isles. Kew Bulletin 47(3): 502. https://doi.org/10.2307/4110576
- Stamatakis A (2014) RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 30(9): 1312–1313. https://doi.org/10.1093/bioinformatics/btu033
- Stielow B, Bubner B, Hensel G, Münzenberger B, Hoffmann P, Klenk HP, Göker M (2010) The neglected hypogeous fungus Hydnotrya bailii Soehner (1959) is a widespread sister taxon of Hydnotrya tulasnei (Berk.) Berk. & Broome (1846). Mycological Progress 9(2): 195–203. https://doi.org/10.1007/s11557-009-0625-1
- Svrček M (1955) Co jest Hydnotrya carnea (Corda) Zobel? Česká Mykologie 9(4): 185–189.
- Tao K, Liu B (1989) Preliminary study on Hydnotrya from China. Journal of Shanxi University (Nat. Sci. Ed. ) 12(1): 81–85.
- Tedersoo L, Hansen K, Perry BA, Kjøller R (2006) Molecular and morphological diversity of pezizalean ectomycorrhiza. The New Phytologist 170(3): 581–596. https://doi.org/10.1111/j.1469-8137.2006.01678.x
- Trappe JM (1969) Comments on Szemere’s “Die Unterirdischen Pilze Des Karpatenbeckens”. Mycologia 61(1): 170–174. https://doi.org/10.1080/00275514.1969.12018711
- Trappe JM (1975) Generic synonyms in the Tuberales. Mycotaxon 2(1): 109–122.
- Trappe JM (1979) The orders, families, and genera of hypogeous Ascomycotina (truffles and their relatives). Mycotaxon 9(1): 297–340.
- Trappe JM, Castellano MA (2000) New sequestrate Ascomycota and Basidiomycota covered by the Northwest Forest Plan. Mycotaxon 75: 153–179.
- Truong C, Mujic AB, Healy R, Kuhar F, Furci G, Torres D, Niskanen T, Sandoval-Leiva PA, Fernández N, Escobar JM, Moretto A, Palfner G, Pfister D, Nouhra E, Swenie R, Sánchez-García M, Matheny PB, Smith ME (2017) How to know the fungi: Combining field inventories and DNA-barcoding to document fungal diversity. The New Phytologist 214(3): 913–919. https://doi.org/10.1111/nph.14509
- Vaidya G, Lohman DJ, Meier R (2011) SequenceMatrix: Concatenation software for the fast assembly of multi‐gene datasets with character set and codon information. Cladistics 27(2): 171–180. https://doi.org/10.1111/j.1096-0031.2010.00329.x
- Vilgalys R, Hester M (1990) Rapid genetic identification and mapping of enzymatically amplified DNA from several Cryptococcus species. Journal of Bacteriology 172(8): 4238–4246. https://doi.org/10.1128/jb.172.8.4238-4246.1990
- Wang XC, Yang ZL, Chen SL, Bau T, Li TH, Li L, Fan L, Zhuang WY (2023) Phylogeny and taxonomic revision of the family Discinaceae (Pezizales, Ascomycota). Microbiology Spectrum 11(3): e00207–e00223. https://doi.org/10.1128/spectrum.00207-23
- White TJ, Bruns T, Lee S, Taylor J (1990) Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In: Innis MA, et al. (Eds) PCR Protocols: A Guide to Methods and Applications. Academic Press, New York, 315–322. https://doi.org/10.1016/B978-0-12-372180-8.50042-1
- Xu AS (2000) Two species of Hydnotrya in Xizang. Mycosystema 19(4): 568–569. https://doi.org/10.1016/S0167-4048(00)07001-2
- Xu YY, Wang YW, Li T, Yan XY, Fan L (2018) DNA analysis reveals rich diversity of Hydnotrya with emphasis on the species found in China. Mycological Progress 17(10): 1123–1137. https://doi.org/10.1007/s11557-018-1425-2
- Zhang BC (1991) Morphology, cytology and taxonomy of Hydnotrya cerebriformis (Pezizales). Mycotaxon 42: 155–162.