MycoKeys 4: 1–8, doi: 10.3897/mycokeys.4.2619
Cylindrocladiella hahajimaensis, a new species of
Cylindrocladiella transferred from
Verticillium
Patrik Inderbitzin 1, Richard M. Bostock 1, Krishna V. Subbarao 1
1 Department of Plant Pathology, University of California Davis, One Shields Avenue, Davis, CA 95616, USA
Abstract
We used phylogenetic analyses based on ribosomal internal transcribed spacer (ITS) and beta-tubulin (TUB) sequences and determined that the correct name of ‘Verticillium hahajimaense’ was Cylindrocladiella hahajimaensis. A closest relative could not be determined, since Cylindrocladiella hahajimaensis clustered within the Cylindrocladiella infestans species complex, a poorly resolved group of taxa. Cylindrocladiella hahajimaensis differed from all other members of the Cylindrocladiella infestans species complex by at least 18 substitutions at the two loci. Morphological characters supported the placement within Cylindrocladiella. In addition to the verticillate conidiophores mentioned in the type description, we found evidence for the presence of penicillate conidiophores. Other differences to the type description included the presence of yellow to brown-pigmented hyphae, and the hyaline instead of pigmented chlamydospores.
Keywords
Ascomycetes, Nectriaceae, taxonomy, phylogenetics
Introduction
The genus Cylindrocladiella comprises a small, monophyletic group of fungi in the Nectriaceae related to Cylindrocladium (Schoch et al. 2000). Cylindrocladium and Cylindrocladiella are morphologically similar, they form two different synanamorphs that resemble Penicillium and Verticillium in terms of branching pattern and phialide arrangement, but may bear long hyphal projections with swollen tips that are referred to as filaments with terminal vesicles. Chlamydospores may also be present (Boesewinkel 1982). Differences between Cylindrocladiella and Cylindrocladium include aspects of conidiophore branching, filament morphology, appearance in culture, and conidia morphology and size (Boesewinkel 1982). The teleomorph of Cylindrocladiella where known, belongs to Nectricladiella (Schoch et al. 2000). Cylindrocladiella comprises ten species that have been isolated from soil as well as plant tissues where they may act as pathogens (van Coller et al. 2005; Zhang and Chi 1996).
Verticillate conidiophores that consist of narrow, elongate phialides arranged in whorls along a main axis, have evolved multiple times as determined by molecular data and phylogenetic analyses. Taxonomic conclusions from these studies have resulted in the traditional genus Verticillium being restricted to a small, monophyletic group of plant pathogens in the Plectosphaerellaceae (Gams et al. 2005; Zare et al. 2007) with the majority of former Verticillium species transferred to other genera, including Lecanicillium (Gams and Zare 2001; Zare and Gams 2001a; Zare and Gams 2008), Pochonia and Haptocillium (Zare and Gams 2001b; Zare et al. 2001), as well as Gibellulopsis and Musicillium (Zare et al. 2007).
During recent studies of Verticillium evolution and taxonomy (Inderbitzin et al. 2011a; Inderbitzin et al. 2011b), we received the ex-holotype culture of ‘Verticillium hahajimaense’ isolated from soil in Japan using cucumber seeds as bait (Watanabe et al. 2001). Phylogenetic analyses showed that ‘Verticillium hahajimaense’ belonged to Cylindrocladiella instead as documented and discussed below.
Methods
Taxon sampling, origins of fungal strains and growth conditions
An ex-holotype culture of ‘Verticillium hahajimaense’ (strain MAFF 238172, PD684) was obtained free of charge from the National Institute of Agrobiological Sciences Genebank (NIAS), Japan, single spore purified and maintained in a glycerol solution at -80°C (Inderbitzin et al. 2011a). For all 42 ingroup taxa used by van Coller et al. (2005), ribosomal internal transcribed spacer sequences (ITS) and beta-tubulin sequences (TUB) were downloaded from GenBank.
DNA extraction, PCR amplification, and DNA sequencing
For complete details see Inderbitzin et al. (2011b). The ITS region was PCR amplified using primers ITS1-F (Gardes and Bruns 1993) and ITS4, and sequenced with ITS5 and ITS4 (White et al. 1990), part of TUB was PCR amplified and sequenced with VTubf2/VTubR (Inderbitzin et al. 2011b).
Phylogenetic analyses
Three different datasets were analyzed, the single locus datasets ITS and TUB using parsimony as implemented in PAUP v.4.0b 10 (Swofford 2002), as well as a the combined ITS plus TUB dataset, using parsimony and likelihood implementations in PAUP, and a Bayesian approach using MRBAYES v3.0b4 (Ronquist and Huelsenbeck 2003). The likelihood and Bayesian analyses used an optimal model of DNA evolution determined using MODELTEST 3.7 (Posada and Crandall 1998). DNA sequences were assembled and aligned in GENEIOUS v4.8.5 (Drummond et al. 2010). For complete details of phylogenetic analyses, see Inderbitzin et al. (2011b).
Microscopy
Morphological observations were from cultures grown on PDA. Microscopy was performed using a Leica DM5000 B microscope (Leica Microsystems CMS GmbH, Wetzlar, Germany), with differential interference contrast (DIC) illumination of specimens mounted in water. Photographs were taken with a Leica DFC310 FX camera, using LEICA APPLICATION SUITE VERSION 3.6.0 software.
Results
DNA sequence data obtained and phylogenetic analyses
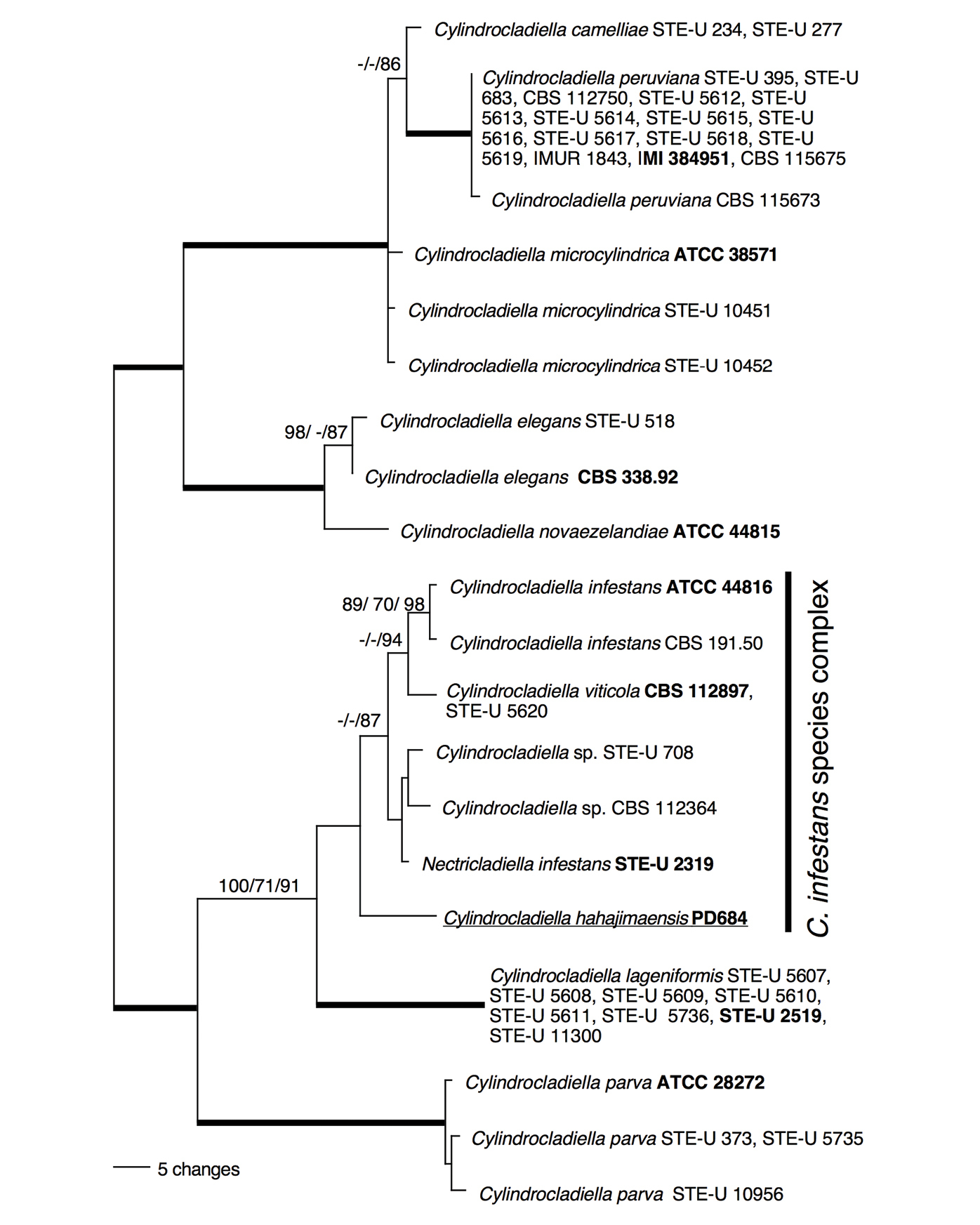
The ITS and TUB sequences of Cylindrocladiella hahajimaensis strain PD684 measured 462 and 480 bp in length, respectively (GenBank JN687561, JN687562), and were aligned with the Cylindrocladiella ingroup sequences by van Coller et al. (2005) resulting in 43 taxa datasets submitted to TreeBase (http://purl.org/phylo/treebase/phylows/study/TB2:S11935). ITS and TUB-derived most parsimonious trees were identical on a 70% bootstrap support level (data not shown), and were combined into one, 950 character dataset that was analyzed using parsimony, likelihood and Bayesian inference of phylogeny, the latter two methods implementing a HKY+I+G model of nucleotide substitution. The 22 most parsimonious trees were 227 steps in length and differed by branches that were supported by less then 70% of the bootstrap replicates (data not shown). One most parsimonious tree was midpoint rooted and is shown in Fig. 1. The most likely tree (-ln likelihood = 2580.47), and the Bayesian tree were congruent with the most parsimonious tree on 70% bootstrap and posterior probability levels, respectively (data not shown, but see Fig. 1 for support values). The tree topology obtained was as in van Coller (2005) for the taxa shared between the two studies, but with lower branch supports probably because our analyses included only two and not three loci. Cylindrocladiella hahajimaensis grouped in a well supported, but poorly resolved clade together with Cylindrocladiella lageniformis, Cylindrocladiella infestans, Nectricladiella infestans, Cylindrocladiella viticola and a Cylindrocladiella sp. (Fig. 1). This group, without Cylindrocladiella lageniformis, was referred to as the ‘Cylindrocladiella infestans species complex’ by van Coller et al (2005). Among its closest relatives, Cylindrocladiella hahajimaensis was most similar to Nectricladiella infestans from which it differed by 18 substitutions in the 950 bp, combined ITS and TUB dataset.
Figure 1.
Phylogenetic position of Cylindrocladiella hahajimaensis within Cylindrocladiella based on a combined ITS and TUB dataset of 950 characters and 43 taxa. The tree is midpoint rooted. One of the most parsimonious trees is shown. Species names are followed by strain identifiers for each strain included in this study, identifiers in bold face represent ex-type cultures. Cylindrocladiella hahajimaensis is underlined. The Cylindrocladiella infestans species complex is delimited by a vertical bar on the right. Numbers by the branches are parsimony, Bayesian and likelihood support values above 70 in that order, branches in bold had maximal support in all analyses.
Taxonomy
Cylindrocladiella
hahajimaensis
(Ts. Watanabe) Inderb., R.M.Bostock and K.V.Subbarao,
comb. nov.
Mycobank: MB483222
Figures 2–4
ºVerticillium hahajimaense Ts. Watanabe, Mycoscience 42: 594 (2001).
Commentary:
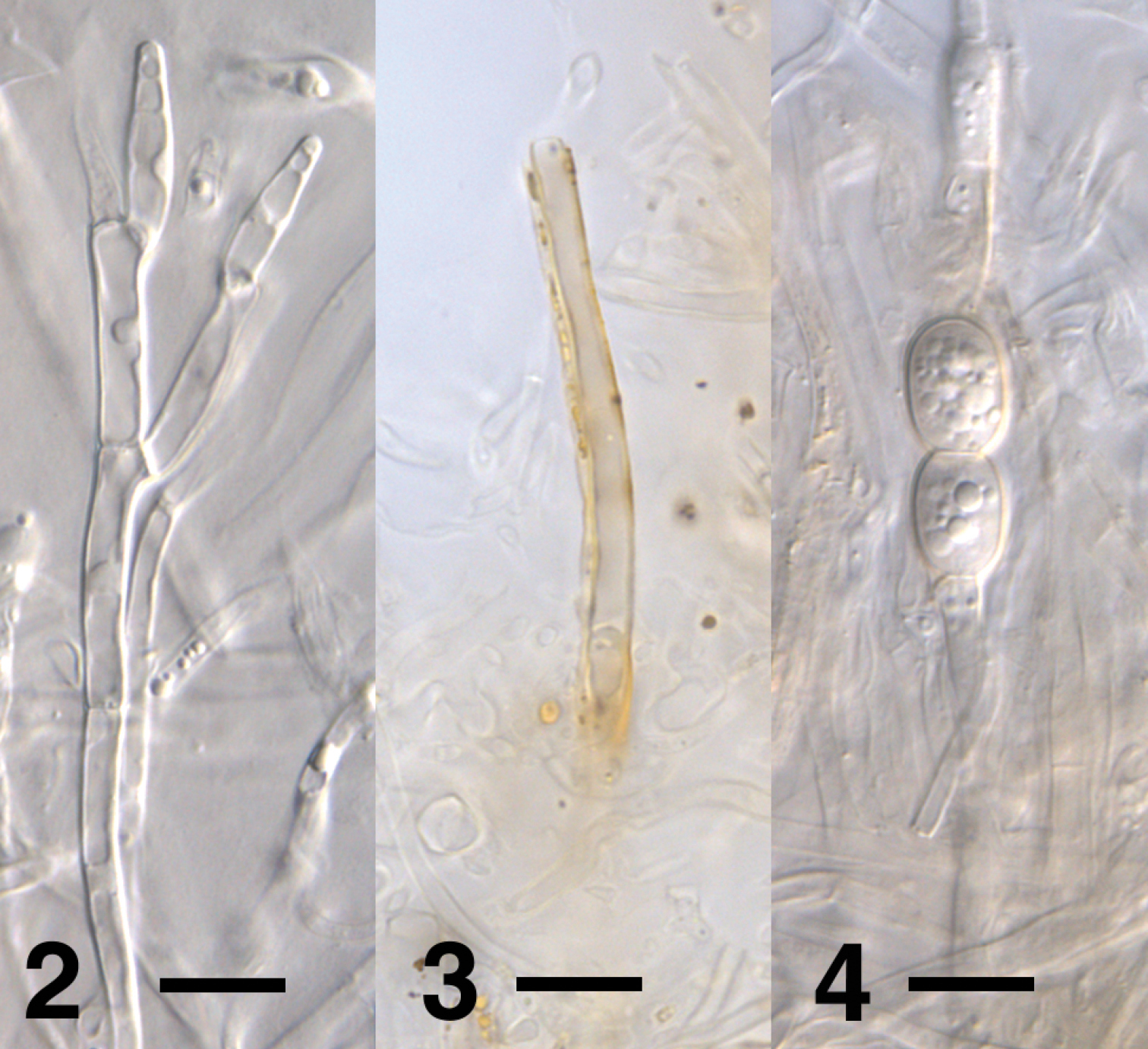
Cylindrocladiella hahajimaensis was formerly placed in Verticillium because of the presence of verticillate conidiophores which were illustrated by Watanabe (2001), together with the chlamydospores and the conidia. We confirmed the presence of these structures, and also found evidence for the presence of penicillate conidiophores (Fig. 2). However, Cylindrocladiella hahajimaensis strain PD684 conidiated sparsely, and we were unable to conclusively assess the morphology of the penicillate conidiophores. In agreement with Watanabe (2001), no filaments with terminal vesicles were seen. Watanabe (2001) described the colony color as ‘Sudan brown or snuff brown’, and the chlamydospores were described as brown. However, we found that the brown colony color was caused primarily by yellow to brown-pigmented hyphae (Fig. 2) not mentioned by Watanabe (2001), the chlamydospores remained hyaline with a brown tinge after 27 days of incubation. We did not find any additional cultures of Cylindrocladiella hahajimaensis at CBS, NIAS or ATCC. More strains of Cylindrocladiella hahajimaensis will need to be examined to update the description of this fungus.
Figures 2–4.
Select morphological features of Cylindrocladiella hahajimaensis strain PD684 (ex-holotype). 2 Potential penicillate conidiophore after eight days on PDA 3 Yellow-pigmented hypha after 15 days on PDA 4 A pair of thick-walled hyaline chlamydospores with brown tinge after 27 days on PDA. Scale bar = 10 µm; Imaging method: DIC.
Cylindrocladiella hahajimaensis is the latest member of Cylindrocladiella which now consists of eleven species (van Coller et al. 2005; Zhang and Chi 1996). Our phylogenetic analyses showed that Cylindrocladiella hahajimaensis was nested within Cylindrocladiella with high support (Fig. 1), and was thus a member of this genus. Morphology did not contradict this placement. The original description of Cylindrocladiella hahajimaensis mentioned verticillate conidiophores and chlamydospores which are present in Cylindrocladiella (Boesewinkel 1982). However, the original description did not mention other characteristics of Cylindrocladiella including penicillate conidiophores and filaments with terminal vesicles. We found evidence for penicillate conidiophores (Fig. 1), but did not find any filaments with terminal vesicles. However, the ex-holotype culture we examined conidiated sparsely, preventing us from corroborating our observations and amending the Cylindrocladiella hahajimaensis type description.
Our analyses included DNA sequence data derived from ex-type cultures of all known Cylindrocladiella species, except for Cylindrocladiella tenuis which has larger conidia than Cylindrocladiella hahajimaensis (Zhang and Chi 1996). Also, none of the species described by Lombard et al. (2012) were included in our analyses. This is because we were unable to replicate the topology of the phylogenetic tree in Lombard et al.’s (2012) figure 1, both based on DNA sequence data retrieved from GenBank, and based on a nexus file with MrBayes block provided by the authors. We do not know the cause of this divergence, but it was consistent and independent of the method of analysis and hardware used. Despite excluding the most recent data available, the analyses we presented here showed strong support that Cylindrocladiella hahajimaensis belonged to Cylindrocladiella, mainly to the Cylindrocladiella infestans species complex (van Coller et al. 2005), a group with uncertain species boundaries. Cylindrocladiella hahajimaensis differed from the most similar member of the Cylindrocladiella infestans species complex included in van Coller et al. (2005) by a considerable 18 nucleotide substitutions at ITS and TUB. More analyses with a larger number of isolates, including the species in Lombard et al. (2012) are needed to resolve the branching order and species boundaries in the Cylindrocladiella infestans species complex, and to determine whether any more recently described species of Cylindrocladiella may be synonyms of Cylindrocladiella hahajimaensis.
Acknowledgements
We would like to thank the National Institute of Agrobiological Sciences Genebank (NIAS), Japan for providing the Cylindrocladiella hahajimaensis culture free of charge and Lorenzo Lombard and Pedro Crous for providing the nexus file. Funding was provided by the California Leafy Greens Board and the United States Department of Agriculture – National Institute of Food and Agriculture (USDA-NIFA).
ReferencesBoesewinkel HJ (1982)
Cylindrocladiella, a new genus to accommodate
Cylindrocladium parvum and other small-spored species of
Cylindrocladium. Canadian Journal of Botany 60: 2288-2294.
Drummond AJ, Ashton B, Buxton S, Cheung M, Cooper A, Heled J, Kearse M, Moir R, Stones-Havas S, Sturrock S, Thierer T, Wilson A (2010) Geneious v4.8.5, Available from
http://www.geneious.com
Gams W, Zare R (2001) A revision of
Verticillium sect.
Prostrata. III. Generic classification. Nova Hedwigia 72: 329-337.
Gams W, Zare R, Summerbell RC (2005) Proposal to conserve the generic name
Verticillium (anamorphic Ascomycetes) with a conserved type. Taxon 54: 179.
Gardes M, Bruns TD (1993) ITS primers with enhanced specificity of basidiomycetes: application to the identification of mycorrhizae and rusts. Molecular Ecology 2: 113-118.
Inderbitzin P, Bostock RM, Davis RM, Usami T, Platt HW, Subbarao KV (2011a) Phylogenetics and taxonomy of the fungal vascular wilt pathogen
Verticillium, with the descriptions of five new species. PLoS ONE 6: e28341.
Inderbitzin P, Davis RM, Bostock RM, Subbarao KV (2011b) The ascomycete
Verticillium longisporum is a hybrid and a plant pathogen with an expanded host range. PLoS ONE 6: e18260.
Lombard L, Shivas R, To-Anun C, Crous P (2012) Phylogeny and taxonomy of the genus
Cylindrocladiella. Mycological Progress: 1–34.
Posada D, Crandall KA (1998) Modeltest: Testing the model of DNA substitution. Bioinformatics 14: 817-818.
Ronquist F, Huelsenbeck JP (2003) MRBAYES 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 19: 1572-1574.
Schoch CL, Crous PW, Wingfield MJ, Wingfield BD (2000) Phylogeny of
Calonectria and selected hypocrealean genera with cylindrical macroconidia. Studies in Mycology 45: 45-62.
Swofford DL (2002) PAUP*. Phylogenetic Analysis Using Parsimony (*and Other Methods). Version 4. Sinauer Associates, Sunderland, Massachusetts, pp.
van Coller GJ, Denman S, Groenewald JZ, Lamprecht SC, Crous PW (2005) Characterisation and pathogenicity of
Cylindrocladiella spp. associated with root and cutting rot symptoms of grapevines in nurseries. Australasian Plant Pathology 34: 489-498.
Watanabe T, Watanabe Y, Fukatsu T (2001) New species of
Acremonium,
Cylindrocarpon and
Verticillium from soil in the Bonin (Ogasawara) Islands, Japan. Mycoscience 42: 1340-3540.
White TJ, Bruns TD, Lee SB, Taylor JW (1990) Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In: Innis MA, Gelfand DH, Sninsky JJ, White TJ (Eds). PCR Protocols. Academic Press, Inc., San Diego: 315-322.
Zare R, Gams W (2001a) A revision of
Verticillium section
Prostrata. IV. The genera
Lecanicillium and
Simplicillium gen. nov. Nova Hedwigia 73: 1-50.
Zare R, Gams W (2001b) A revision of
Verticillium section
Prostrata. VI. The genus
Haptocillium. Nova Hedwigia 73: 271-292.
Zare R, Gams W (2008) A revision of the
Verticillium fungicola species complex and its affinity with the genus
Lecanicillium. Mycological Research 112: 811-824.
Zare R, Gams W, Evans HC (2001) A revision of
Verticillium section
Prostrata. V. The genus
Pochonia, with notes on
Rotiferophthora. Nova Hedwigia 73: 51-86.
Zare R, Gams W, Starink-Willemse M, Summerbell RC (2007)
Gibellulopsis, a suitable genus for
Verticillium nigrescens, and
Musicillium, a new genus for
V. theobromae. Nova Hedwigia 85: 463-489.
Zhang C, Chi P (1996) A new species of
Cylindrocladiella, an unrecorded genus from China. Acta Mycologica Sinica 15: 170-172.